Most literalist creationists argue that there are no transitional forms (e.g. Morris 1967, Gish 1979, 1985, 1995). Cracraft (1983) suggests that this argument,
could well be the paradigm example characterizing the entire creation-evolution controversy for it illustrates how creationists have taken an extremely complex scientific question, simplified the matter to the point of misrepresentation, and then have promoted the blatantly false claim that the fossil record supports the creationist world view. Indeed, from the manner in which creationists have discussed this issue, one can only conclude that either the creationists have consciously adopted the tactic of outright distortion or they are so abysmally ignorant of the scientific arguments and data that their apparent distortions are only accidental, not purposeful. (p. 178)
Cracraft also discusses the methodology employed by creationists on this topic.
Creationists have adopted three lines of argumentation against the existence of transitional forms: (1) they quote liberally from various paleontologists as to the paucity of transitional forms; (2) they define the concept of "transitional form" in a way that is distinctly different from the evolutionists' use of the term; and (3) they simply deny the existence of intermediate taxa, whilst ignoring the vast scientific literature opposing their position. (p. 179).
In discussions about the existence of transitional forms in the fossil record, no fossil has caused more feathers to fly than Archaeopteryx, due to its pride of place as a classic example of a transitional form - in evolutionary terms, a form which exhibits characters shared with one group and only that group, whilst also exhibiting other characters shared with another group and only with that group (e.g. Kitcher 1982), in other words a morphological intermediate. Dr. Duane Gish of the Institute for Creation Research, is probably one of those most vocal in crying foul at the recognition of Archaeopteryxas a transitional form (e.g. Gish 1979, 1985, 1995). There have been various commentaries on passages from Dr. Gish's 1978 book Evolution? The Fossils Say No! (e.g. Kitcher 1982; Cracraft 1983; Raup 1983; Halstead 1984; Strahler 1987; Blackburn 1995), but relatively little commentary about Dr. Gish's 1985 book Evolution: The Challenge of the Fossil Record. This article looks at a small portion of the 1985 book - that pertaining to Archaeopteryx - in light of Cracraft's comments.
Dr. Gish has recently (1995) published an updated version of his 1985 book, called, Evolution: The fossils still say NO!. An analysis of the treatment of Archaeopteryx in that book is underway. However, the well known half life of literalist creationist material, long after it has been superceded, means that this may still represent a valuable resource for people replying to literalist creationist statements about Archaeopteryx.
Some material from Evolution: Challenge of the Fossil Record has been rendered obsolete by new data accrued in the 10 years since the book was first published. However...
"In reference to Archaeopteryx, Ichthyornis, and Hesperornis, Beddard stated: "So emphatically were all these creatures birds that the actual origin of Aves is barely hinted at in the structure of these remarkable remains" (Beddard 1898, p. 160)." (p. 110)
Despite the fact that Beddard was writing in 1898, when only two specimens of Archaeopteryx were known, Beddard's (1898, p. 6) criterion for identifying birds is instructive. "A bird may be known by its feathers; to define a bird it is only necessary to refer to its covering of feathers. No other animal has any structures comparable to a well developed feather." Note however that Beddard was in no doubt of the uniqueness of Archaeopteryx, listing some 8 characters in which "...Archaeopteryx differed from all known birds." (p. 532). It is interesting to note here that the recent dicovery of Sinosauropteryx - a dinosaur in China which appears to preserve feathers - suggests that, at least as far as fossil species are concerned, feathers may not be such a good defining character for birds
These, characters are shared with reptiles and have become more refined with the finding of additional specimens, especially the Eichstätt specimen, described in 1974. Such were the differences between Archaeopteryx and all other birds that Beddard (1898, p. 159) was prompted to write, "[t]here can be no question, in my opinion, that birds must be primarily divided into two great divisions, viz. Saururae and Ornithurae, the first contains Archaeopteryx and possibly Laopteryx, the latter the rest of birds, both living and extinct." Clearly, although Beddard considered Archaeopteryx a bird because it possessed feathers, he considered it sufficiently different to all other birds to merit its own division (Laopteryx was later reclassified as a pterosaur by Ostrom in 1986).
"During the eighty five years since publication of Beddard's book, no better candidate as an intermediate between reptiles and birds than Archaeopteryx has appeared. Not a single intermediate with part-way wings or part-way feathers has been discovered." (p. 110)
This is an example of defining a transitional form in such a way as to eliminate the possibility of ever finding one (Cracraft 1983), since such "part-way" intermediates would not be expected. The evolution of features does not occur at the same time nor at the same rate. Some characters evolve rapidly, others more slowly, so that smooth, nicely "part-way" intermediates will not be found. During the last eighty five years, mosaic forms intermediate between Archaeopteryx and birds, and between Archaeopteryx and reptiles have been found. For example, Mononychus (Altangerel et al. 1993), "provides a character set of great importance to understanding primitive birds, this set can be interpreted unambiguously and it indicates a transitional position between Archaeopteryx and all other birds." (Milner 1993, p. 589). Also, the finding of Sinosauropteryx suggestd that feathers may have come first and that feathered dinosaurs evolved into birds. Thus an intermediate between dinosaurs and birds would already possess feathers.
"Not a single intermediate with part-way wings or part-way feathers has been discovered. Perhaps this is why, with the passage of time Archaeopteryx in the eyes of some evolutionists, has become more and more 'reptile-like'! In contrast to Beddard's assessment of Archaeopteryx , some evolutionists today not only assert that this bird is undoubtedly linked to reptiles but that if clear impressions of feathers had not been found, Archaeopteryx would have been classified as a reptile. This is a gross overstatement to say the least." (p. 110-111)
The "gross overstatement" is Dr. Gish's, on several grounds.
On the link to reptiles, Beddard (1898, p. 154) states, "the general belief is in the origin of birds from some reptile stem, but there is not an absolute agreement as to precisely which group of reptiles birds are most nearly akin to. The researches of Marsh and Huxley, besides those of Cope, Seeley, Hulke and others, have led to a general acceptance of a nearer kinship with the dinosaurs than with any other group of reptiles." Also, "[i]t follows, therefore, that in sketching, at any rate, the main outlines of our scheme attention must be paid only, or chiefly, to those characters which birds have inherited from their reptilian ancestors." (Beddard 1898, p. 159). It is clear that in Beddard's view, Archaeopteryx was linked to reptiles and thus there is no "contrast" between his views and the modern view.
The reptilian character of Archaeopteryx has long been noted. In 1867, Huxley described birds as being greatly modified reptiles (Newton 1884). Indeed, the very author and book that Dr. Gish cites (Beddard 1898, p. 164) stated, "[t]here can be no doubt that the Archaeopteryx, far though it may have diverged from the ancestral stock, has retained more of the reptile than any other form known to us." Jordan & Kellogg (1916, p. 301) stated that, "A comparison of the ancient reptiles with the long-tailed Archaeopteryx and other toothed birds shows that the birds and reptiles were once scarcely distinguishable, although now so very different. Birds have feathers, reptiles do not; but there is scarcely any other permanent difference." "We may now stop talking about 'the missing link' between birds and reptiles. So much so is Archaeornis [the Berlin specimen of Archaeopteryx] this link that we may term it a warm-blooded reptile disguised as a bird." (Heilmann 1926, p.32). "Archaeopteryx, on the other hand, is nearly one hundred per cent reptilian, and if it were not for the feathers no-one would hesitate to regard it as purely reptilian or even dinosaurian." (Lowe 1935, p. 408). "There is in my opinion, nothing in the entire skeleton which could be pointed to as being definitely avian as opposed to dinosaurian." (Lowe 1935, p. 409). "[T]he close relationship of reptiles and birds is generally accepted and is a matter of common knowledge and history." (Lowe 1935, p. 399). "Archaeopteryx and Archaeornis then, though indubitably birds, as witnessed by their well developed and typical flight feathers, their limbs and indeed their whole skeletons, proclaim their reptilian ancestry in no uncertain fashion." (Tucker 1938, p. 322-323).
The above quotes (see also the Lull quote below) show that Dr. Gish's claims are false. Neither the reptilian affinities of Archaeopteryx, nor the statements that without feathers Archaeopteryx would be classified as a reptile, are recent constructs.
"From the reconstruction shown in Figure 11, it is obvious that Archaeopteryx was very much a bird, equipped with a bird-like skull, perching feet, wings, feathers and a furcula, or wish-bone." (p 112).

|
The figure (reprinted here as Fig. 1) is taken from Lull (1940, pl. XIV). However, the caption to plate XIV cites Heilmann as the source. Dr. Gish's Figure 11 is in fact, a painting, by Heilmann, from the frontispiece of his 1926 book, The Origin of Birds. Lull (1940, p. 328) describes the figure, "[t]hese first birds, of which but two or three specimens have been recovered, are known as Archaeopteryx and Archaeornis (see Fig. 80 and Pl. XIV) are so reptile-like that were it not for the preserved feathers it is doubtful whether they could be surely proved to be birds." Prophetic words, since in 1940 a specimen of Archaeopteryx lay in the Haarlem Museum, misidentified as a pterosaur, and more recently, the Solnhofen specimen was recognised as being an Archaeopteryx after being originally identified as the small dinosaur Compsognathus.
Dr. Gish not only ignores Lull's clear proclamation concerning the reptilian nature of Archaeopteryx, but uses a 70 year old painting of Archaeopteryx, drawn as a bird, to show that Archaeopteryx was "very much a bird". Presumably, by the same logic, "Barney" proves that dinosaurs were purple!
The painting does not show a "bird-like skull"; the feet are obscured by the foliage; and the furcula is an internal skeletal structure, thus making its exhibition in the painting impossible. Besides, in Heilmann's (1926) detailed analysis, Archaeopteryx was compared with maniraptoral dinosaurs only for the link to be rejected because it was thought that maniraptoral dinosaurs lacked clavicles (these are thought to be the precursor to the avian furcula or wishbone) (Ostrom 1976). More recently, not only have clavicles been found in dinosaurs such as Velociraptor, Euparkeria and Ornithosuchus (Bryant & Russell 1993), but indeed furculas have also been found in dinosaurs such as Oviraptor and Ingenia (Barsbold 1983, Barsbold et al. 1990, Bryant & Russell 1993). Therefore, possession of a furcula is no longer a character unique to birds.
The wings of Archaeopteryx are structurally dissimilar to those of modern birds; the wrist and finger bones are unfused (as in most theropod dinosaurs, but unlike birds and some cretaceous dinosaurs, in which the bones are fused); the wrist articulation is also much less than that in modern birds; the fingers retain claws in the adult stage (as in other theropod dinosaurs but unlike birds) and the shoulder joint is most similar to that of the theropod dinosaur Deinonychus and appears intermediate between theropod dinosaurs and birds (Jenkins 1993). Not only does the painting fail to document some of the characters claimed by Dr. Gish, but the painting is patently inadequate as a scientific illustration - since it was never meant to be one. A far more accurate figure to illustrate the true nature of Archaeopteryx is shown in Figure 2.
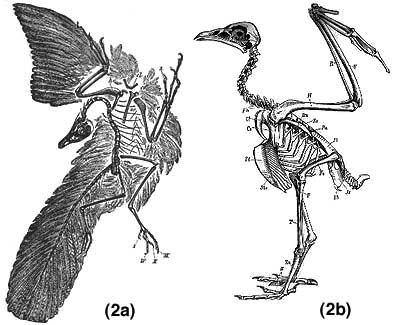
|
This comparison between Archaeopteryx (a) (Parker & Haswell 1940) and a modern bird (b) (Claus 1889) shows the skeletal structure in detail and highlights the differences. This figure clearly shows that the wing and skull of Archaeopteryx differs from that of modern birds.
"It has been claimed that the skull of Archaeopteryx was reptile-like rather than bird-like. Recently, however, the cranium of the 'London' specimen has been removed from its limestone slab by Whetstone (1983). Studies have shown that the skull is much broader and more bird-like than previously thought (Whybrow 1982)." (p. 112-113)
There appears to be some confusion here, since the cranium of the London specimen was prepared out by P.G. Whybrow in 1980 and the problems encountered, the tools and conservation methods used during the preparation were detailed in a technical article (Whybrow 1982), Whetstone (1983) described and interpreted the newly exposed cranium, not its removal. It appears that Benton (1983) (the reference cited by Dr. Gish in his next sentence - see below) incorrectly attributed the description of the skull to Whybrow. Whybrow actually makes no comment on the bird-like nature of the skull. The error appears to have been repeated by Dr. Gish, who also incorrectly cites Whybrow as the source of this statement. By citing Whetstone and Whybrow Dr. Gish gives the impression that the primary literature had been consulted when, in fact, it appears not to have been. The "bird-like" comparison comes from Whetstone , "[t]he skull is much broader and more bird-like that earlier interpreted by de Beer (1954), supporting the estimates of brain size by Jerison (1973)." (Whetstone 1983, p. 439).
The original quote is somewhat different from "...the skull is much broader and more bird-like than previously thought", since Whetstone compares the skull with de Beer's interpretation and indicated that a larger skull size had already been suggested by Jerison in 1973. Some clarification might be in order at this point. In his description of Archaeopteryx, in 1954, de Beer thought that the skull was exposed along the mid-line, suggesting that half of the braincase was exposed and half buried in the matrix. This allowed de Beer to make an estimate of the brain size. However, as Jerison (1973) pointed out, de Beer's interpretation of the mid-line was in error, and only approximately one third of the braincase was exposed. Therefore de Beer's reconstruction underestimated the brain size. Excavation of the cranium by Whybrow confirmed Jerison's interpretation. An enlarged cranium relative to the rest of the skull is a character of birds, in reptiles the cranium is usually smaller. However, an enlarged cranidium is not unique to birds. Several dinosaurs also have enlarged craniums, including Sauronrnithoides (Hopson 1980), Stenonychosaurus (Currie 1985) and Troodon (Currie & Zhao 1993). In some cases the dinosaur cranium is more bird-like than the cranium of Archaeopteryx.
Whetstone does describe the braincase (as opposed to the skull) of Archaeopteryx as, "typically avian" (p.449), however, he also describes skull features found in Archaeopteryx and not in birds.
"This has led Benton to state that 'Details of the braincase and associated bones at the back of the skull seem to suggest that Archaeopteryx is not the ancestral bird ... (Benton 1983).' Benton can only suggest that Archaeopteryx may be an offshoot from the early avian stem." (p. 112-113).
To reconstruct the original quote: "Details of the braincase and associated bones at the back of the skull seem to suggest that Archaeopteryx is not the ancestral bird, but an offshoot from the early avian stem." (Benton 1983, p. 99). However, whether or not Archaeopteryx is the ancestral bird does not detract from the fact that is a transitional form. Transitional because it possesses characteristics in common with reptiles and also characteristics in common with birds. In other words, it possesses a mosaic of characters, some obviously reptilian (such as the long bony tail with many free vertebrae and the sacrum with 6 vertebrae; some transitional, such as the pelvic girdle and the length of the arms; and some bird-like, such as feathers and an opposable big toe). In fact, a recent study of the new, seventh, Archaeopteryx specimen by Elzanowski & Wellnhofer (1996) has highlighted this mozaic nature by documenting that the skull has avian (palatine + maxillary, hook shaped choanal process with a long pterygoid wing) and theropod (single vomer, and hook shaped jugal process of the ectopterygoid) traits. The transitional nature of Archaeopteryx is due to its morphology, not its taxonomy.
"John Ostrom has of late been the foremost proponent of a dinosaurian ancestry for Archaeopteryx. However, Tarsitano and Hecht have criticized Ostrom's hypothesis, claiming amongst other things that he had misrepresented the homologies of the limbs of Archaeopteryx and theropod dinosaurs (Tarsitano & Hecht 1980). (p. 113).
This appears to have been taken from the Benton (1983) reference noted above, suggesting that Benton, and not Tarsitano & Hecht 1980, was Dr. Gish's primary source. If this was the case, Dr. Gish has omitted the next line. The full statement from Benton, (compare with Dr. Gish's statement above) reads, "Finally, Tarsitano and Hecht (1980) criticized various aspects of Ostrom's hypothesis, and they considered that he had misrepresented the homologies of the limbs of Archaeopteryx and theropods. Thulborn & Hamley (1982), reviewing all the criticisms [of Tarsitano and Hecht and of Martin et al. 1980 - see below], however, have concluded that they are without foundation (incorrect interpretations, inconclusive evidence, persistence of primitive characters), and that they do not "seriously weaken the hypothesis that Archaeopteryx is closely related to theropod dinosaurs"." (p. 99-100). Dr. Gish appears to have ignored dissenting commentary on Tarsitano & Hecht 1980 to strengthen his arguement.
"Martin, Stewart and Whetstone have also challenged Ostrom's claim that birds were derived from dinosaurs (Martin et al. 1980). Their analysis centers on the structure of the avian tarsus (ankle) and avian teeth. They say, 'Ostrom...has stated that the skeleton of Archaeopteryx is essentially identical with that of some small theropod dinosaurs' ... We think that many of these 'coelurosaurian' features are incorrectly identified. This is certainly true of the tarsal region, where Archaeopteryx has a pretibial bone, fibula and calcaneum of the avian type. In the dentition, Archaeopteryx has unserrated teeth with constricted bases and expanded roots like those of other Mesozoic birds." (p. 113)
In their 1980 paper, Martin, Stewart and Whetstone supported the theory that crocodiles were more closely related to birds than any other group. However their paper has been criticized by a number of other palaeontologists. Both Thulborn & Hamley (1982) and Cracraft (1986) commented that the analyses of Martin et al. (amongst others) were either insufficiently comparative, or that the arguments presented were not supported by rigorous phylogenetic methods. More specific criticism comes from Howgate (1984, p. 173) who, in reply to the claims made for the teeth of Archaeopteryx, states that, "no matter how similar are the teeth of Cretaceous birds and of crocodiles, there is little similarity of either to the teeth of Archaeopteryx. Of the characters supposedly indicative of close relationships only one is present, namely the lack of serrations." Thulborn & Hamley (1982, p. 623) comment that, "Martin et al. (1980, p. 89) have made a similar suggestion, maintaining that the ascending process of the Archaeopteryx ankle is 'primarily associated with the calcaneum'. However, these authors present a decidedly ambiguous reconstruction of the Archaeopteryx ankle; it shows the ascending process associated equally with the astragalus and calcaneum. The Archaeopteryx ankle depicted by Martin et al. (1990, fig. 1G) seems, in fact, to be structurally intermediate between the theropod ankle and the neornithiform bird ankle." By 1985, the leading proponent of the crocodilian theory, Walker, had decided that the theory was no longer tenable (Dobson 1985).
Although there was disagreement over which reptile group were ancestral to birds, all these workers agreed that birds were descended from a group of reptiles.
"The presence of claws on the wings of Archaeopteryx are often cited as evidence of a reptilian ancestry. However, there are at least three birds very much alive and well today that have claws on the wings, but no one for a moment would claim that any of these is intermediate between reptile and bird. The hoatzin (Opisthocomus hoatzin), a South American bird, possesses two claws in its juvenile stage. Furthermore, it is a poor flyer, with an astonishingly small keel, another feature attributed to Archaeopteryx. The young of the touraco (Touraco corythaix) an African bird, has claws on each wing and the adult is also a poor flyer. The ostrich has three claws on each wing, which, if one chose to do so, could be characterized as even more reptile-like than those of Archaeopteryx." (p. 113-114)
It is significant that claws are present on the fingers of Archaeopteryx, in the adult form. The hoatzin and the touraco lose the claws by the time they reach the adult condition. The claws are present in the juvenile to assist in climbing the dense foliage in which these birds live. In fact almost all birds exhibit claws tosome extent, usually in the embryonic stage (e.g. Romanoff 1960), but they are lost by the time the bird hatches. As McGowan (1984, p 123) says, "in retaining a primitive reptilian feature which other birds lose just before leaving the egg [the hoatzin] is showing us its reptilian pedigree. Far from being evidence to the contrary, the hoatzin is additional evidence for the reptilian ancestry of birds."
The shoulder joint of Archaeopteryx is more similar to the theropod dinosaur Deinonychus, than to modern birds (Jenkins 1993), the shoulder joint of the ostrich is more similar to other birds than than to the shoulder joint of Archaeopteryx. The wrist joint of the ostrich is partially fused as in modern birds, whereas the wrist joint of Archaeopteryx is not, as in typical reptiles. The fingers of the ostrich wing are fused together as in modern birds, the fingers of Archaeopteryx are free, as in typical reptiles. Thus, the wing of the ostrich can in no way be described as "even more reptile-like than those of Archaeopteryx."
"Another alleged reptilian feature of Archaeopteryx was its possession of teeth. If this is a feature derived from a reptilian ancestor, and toothed birds subsequently evolved into toothless birds, then the fossil record should produce intermediates documenting the gradual loss of teeth in birds. Not one single intermediate has ever been discovered. Some fossil birds have teeth some did not. That this should be true is not surprising since this is true of all other classes of vertebrates - fish, amphibians, reptiles and mammals. Furthermore, following the notion that the absence of teeth denotes a more 'advanced' state, then the duck-billed platypus and the spiny anteater, mammals that do not have teeth, should be considered more advanced or highly evolved than man, yet in many other ways, as previously mentioned, the duck-billed platypus and spiny anteater could be considered the most primitive of all mammals. Thus, the possession or absence of teeth proves nothing about ultimate ancestry." (p. 114)
The lack of intermediates showing tooth reduction has more to do with the lack of fossils and the way evolution operates than with any lack of such intermediates in the history of the birds. The number of Jurassic and Cretaceous bird fossils number only a few dozen at best. It is not surprising that such intermediates are not represented. However, as has been pointed out previously the expectation of finding such smooth intermediates is fallacious.
"Advanced" and "primitive" carry certain connotations. Biologists now use 'derived' for "advanced" and 'ancestral' for "primitive". However, what are considered derived characters for one group cannot be used to decide what are derived characters in another group. For instance, derived characters in snakes are (in order of appearence); reduction of limbs, loss of limbs and reduction from two to one lung, acquisition of fangs, acquisition of sensory pits. Thus from the point of view of snake evolution, humans, with their retention of limbs, retention of two lungs, absence of fangs and absence of sensory pits, are ancestral, or "primitive". As can be seen from this example what is sauce for the snake is certainly not sauce for the human! The absence of teeth is considered a derived character in birds, it is of little importance when deciding which characters are "advanced" in other groups. Thus the claim that, "the duck-billed platypus and the spiny anteater, mammals that do not have teeth, should be considered more advanced or highly evolved than man" is ludicrous, and Dr Gish should know this.
"Evolutionists have long maintained that contemporaries could not have an ancestral-descendant relationship but if related, they must have evolved from a common ancestor sometime in the past." (p. 116)
This is only true for populations, not for species in general. A population can be described as a (usually) reproductively isolated group of individuals which comprise a specific gene pool. Changes to that gene pool over time (the death of individuals, the birth of new individuals, mutations, etc.) all conspire to alter the genetic composition of the population so that the descendant population represents the original population plus the sum of the changes over time. Thus the descendant population cannot coexist with the ancestral population because the descendant population is the ancestral population, plus the sum of the changes over time. However, in the vast majority of cases, one population does not comprise the entire species. There are usually multiple populations spread out geographically, some interact, some do not. What happens in one isolated population is not mirrored in the species as a whole. Indeed, changes in isolated populations is one of the engines powering speciation. Whilst the descendant population cannot be contemporaneous with its ancestral population, it can be contemporaneous with the ancestral species. A good example is the polar bear, Ursus maritimus.
It is thought that polar bears evolved from a population of brown bears (Ursus arctos) which became isolated above the Arctic Circle during one of the last major glacials. Separated from the main populations of brown bears, which remained further south, this ancestral population adapted to the cold climate (fur on the feet, white coat for camouflage during hunting, etc.), evolving into Ursus maritimus, whilst the more southerly populations retained the original characters. In this example, the modern polar bears cannot coexist with the ancestral population of brown bears from which they are considered to have descended, because they are that population, plus the sum of the genetic changes that have occurred over time. However, they can, and obviously do, coexist with the ancestral species - Ursus arctos. What Dr. Gish is trying to suggest, is that two contemporaneous species cannot have an ancestor-descendant relationship. As the polar bear example shows, this is not the case. Indeed ancestral species have been known to outlive their descendant species (e.g. Ozawa 1975).
"In a somewhat similar fashion, Archaeopteryx, although unquestionably a bird, was a mosaic which included some features that are usually termed 'reptilian.' In this respect, it is interesting to note the comment of Steven Jay Gould of Harvard University and Niles Eldredge of the American Museum of Natural History, both Ardent anticreationists. They state that, 'At the higher level of evolutionary transition between basic morphological designs, gradualism has always been in trouble, though it remains the "official" position of most Western evolutionists. Smooth intermediates between Baupläne are almost impossible to construct, even in thought experiments; there is certainly no evidence for them in the fossil record (curious mosaics like Archaeopteryx do not count)' (Gould & Eldredge 1977, p. 147). There are several important aspects of this statement, each of which seriously damages the credibility of evolutionary theory." (p.114-115)
There certainly are important aspects to the statement of Eldredge and Gould, as used by Dr. Gish, but the damage is to the credibility of creationists. The article in question is a discussion of punctuated equilibrium and how the evidence from the fossil record does not support a purely gradualistic evolutionary model. Eldredge and Gould point out that intermediates, for the most part, do not exhibit such smooth evolution, but that features evolve at differing rates. In this they were following the ideas put forward by de Beer (1969, p. 133-134; first published as de Beer 1954), who suggested that, "the statement that the animal was intermediate might mean that it was a mixture and that the transition affected some parts of the animal and not others, with the result that some parts were similar to those of one type, other parts similar to another type, and few or no parts intermediate in structure. In such a case the animal might be regarded as a mosaic in which the pieces could be replaced independently one by one, so that the transitional stages were a jumble of characters, some of them similar to those of the class from which the animal evolved, others similar to those of the class in which the animal was evolving. If now it be asked what kind of transition is shown by Archaeopteryx, the answer is perfectly clear. It is a mosaic in which some characters are perfectly reptilian and others no less perfectly avian."
Thus being a "mosaic" does not disqualify a form from being an intermediate, merely a smooth intermediate.
"Not only is it impossible at this level to find a smooth series of intermediates in the fossil record, it is impossible to imagine what such intermediates may have looked like (for example, try to imagine am emergent Pteranodon with half a jaw and half a wing!). Finally, note that Gould and Eldredge specifically exclude Archaeopteryx as a transitional form, terming it, as is the duck-billed platypus, a strange mosaic that doesn't count. So much for Archaeopteryx as an intermediate!" (p.115)
This represents a clear misrepresentation of Gould and Eldredge This probably ranks as the most famous case of the misrepresentation of scientists by a creationist author. From the previous discussion, it is obvious that Eldredge and Gould did not "specifically exclude Archaeopteryx as a transitional form", but merely as an example of a smooth transitional form. This kind of misrepresentation has led Gould to clarify his position. On transitional forms in general, he states, "since we proposed punctuated equilibrium to explain trends, it is infuriating to be quoted again and again by creationists - whether through design or stupidity, I do not know - as admitting that the fossil record includes no transitional forms. Transitional forms are generally lacking at the species level but are abundant between larger groups." (Gould 1983, p. 260). More specifically, Gould (1991, p. 144-145) states that "Archaeopteryx, the first bird, is as pretty an intermediate as paleontology could ever hope to find." Strange words from someone who "specifically exclude[s] Archaeopteryx as an intermediate"!
"Swinton, an evolutionist and an expert on birds, states: The origin of birds is largely a matter of deduction. There is no fossil evidence of the stages through which the remarkable change from reptile to bird was achieved (Swinton 1960). Romer has said that: This Jurassic bird [Archaeopteryx] stands in splendid isolation; we know no more of its presumed thecodont ancestry nor of its relation to later 'proper' birds than before (Romer 1968)." (p. 114)
Both Swinton and Romer were writing some 16 and 8 years, respectively, before Ostrom's (1976) seminal work on the relationships between Archaeopteryx and maniraptoral dinosaurs. Their comments are no longer representative of current thinking on the origin of birds, nor the ancestry of Archaeopteryx, and were not for some 9 years before Dr. Gish wrote his book.
"A recent discovery by paleontologist James Jensen has dealt an especially serious blow to the claim that Archaeopteryx represents a transitional form between reptiles and birds. Jensen has found what he believes to be fossil remains of undoubted modern birds in rocks of the Upper Jurassic, the rocks in which Archaeopteryx has been found. Regardless of what one believes about the time scale or the geological column, this discovery, if ultimately verified, means that Archaeopteryx was a contemporary of modern birds." (p. 116)
This refers to an article by Jensen in 1981, where several avian-like bones were described, with the proximal part of a tibiotarsus were given the name Palaeopteryx thomsoni. However, Jensen & Padian (1989) re-identified this bone as belonging to the theropod dinosaur Deinonychus. Their conclusions were deliberately blunt : "No material described here is unquestionably avian. Most is pterodactyloid. Several specimens pertain to the monophyletic group formed by birds and deinonychosaurs. Archaeopteryx is the earliest known bird; these Morrison Formation sediments are younger that the Solnhofen limestones from which Archaeopteryx comes." (p. 372)
Dr. Gish states that Archaeopteryx was, "very much a bird" (p. 112), and, "an undoubted true bird" (p. 116). However, the classification of Archaeopteryx as a bird says nothing about its ancestry. As Raup (1983, p.157) says, "[t]he practicing paleontologist is obliged to place any newly found fossil in the Linnean system of taxonomy. Thus, if one finds a birdlike reptile or a reptilelike bird (such as Archaeopteryx), there is no procedure in the taxonomic system for labeling and classifying this as an intermediate between the two classes Aves and Reptilia. Rather, the practicing paleontologist must decide to place his fossil in one category or the other. The impossibility of officially recognising transitionary forms produces an artificial dichotomy between biologic groups. It is conventional to classify Archaeopteryx as a bird. I have no doubt, however, that if it were permissible under the rules of taxonomy to put Archaeopteryx in some sort of category intermediate between birds and reptiles that we would indeed do that."
Many of Dr. Gish's assertions about Archaeopteryx have been shown to be incorrect. Some of these claims have been rendered invalid due to evidence found since Dr. Gish's book was first published. However, in a significant number of cases, invalidating evidence was known at the time the claims were made.
Acknowledgements
I would like to thank Rich Trott for initially suggesting this project and supplying valuable material and advice. Marianne Cotugno is thanked for a valuable review of the first draft and Tom Holtz is thanked for suggesting the polar bear example. The text was originally coverted to HTML format for the talk.origins archive by Brett Vickers.
This is a University of Ediacara Palaeontological Contribution.
References
Altangerel, P.; Norell, M. A.; Chiappe, L. M. & Clark, J. M. 1993. Flightless bird from the Cretaceous of Mongolia. Nature, 362: 623-626
Barsbold, R. 1983. [Carnivorous dinosaurs from the Cretaceous of Mongolia]. Trudy Soumestnaya Sovetsko-Mongol'skaya Paleontogicheskaya Ekspeditsiya, 19: 1-117. [In Russan].
Barsbold, R.; Maryanska, T. & Osmolska, H. 1990. Oviraptorosauria. In: Weishampel, D. B.; Dodson, P. & Osmolska, H. (eds.), The Dinosauria. 249-258. University of California Press, Berkeley.
Beddard, F. E. 1898. Structure and Function of Birds. Longmans, Green & Co., London. 548 pp.
Benton, J. 1983. No consensus on Archaeopteryx. Nature, 305: 99-100.
Blackburn, D. G. 1995. Paleontology meets the creationist challenge. Creation/Evolution, 36: 26-38.
Bryant, H.N. & Russell, A.P. 1993. The occurrence of clavicles within dinosauria: implications for the homology of the avian furcula and the utility of negative evidence. Journal of Vertebrate Paleontology, 13:171-184.
Claus, C. 1889. Elementary Text-Book of Zoology. Sonnenschein & Co, London. 352 pp.
Cracraft, J. 1983. Systematics, comparative Biology, and the case against creationism. p. 163-192. In: Godfrey L.R. (ed.), Scientist Confront Creationism. Norton & Co, New York.
Cracraft, J. 1986. The origin and early diversification of birds. Paleobiology, 12: 383-399. Colbert, E. H. & Morales, M. 1991. Evolution of the vertebrates : a history of the backboned animals through time. 4th ed. Wiley-Liss, New York. 470 p.
Currie, P. J. 1985. Cranial anatomy of Stenonychosaurus inequalis (Saurischia, Theropoda) and its bearing on the origin of birds. Canadian journal of Earth Science, 22: 1643-1658.
Currie, P. J. & Zhao, Xi-Jin 1993. A new troodontid (Dinosauria, Theropoda) braincase from the Dinosaur Park Formation (Campanian) of Alberta. Canadian Journal of Earth Science, 30: 2231-2247.
de Beer, G. A. 1954. Presidential Address to the Zoology Section of the British Association for the Advancement of Science, Oxford, Sept. 2nd 1954. The Advancement of Science, 15: no. 42.
de Beer, G. A. 1969. Streams of Culture. J.B. Lippincort, Philadelphia. 237 pp.
Dodson, P. 1985. International Archaeopteryx Conference. Journal of Vertebrate Paleontology, 5(2):177
Elzanowski, A. 1996. Crainial morphology of Archaeopteryx: Evidence from the seventh skeleton. Journal of Vertebrate Paleontology, 16: 81-94.
Gish, D. T. 1979. Evolution? The fossils say No! Creation-Life, San Diego. 186 pp.
Gish, D. T. 1985. Evolution: Challenge of the Fossil Record. Creation-Life, San Diego. 278 pp.
Gish, D. T. 1995. Evolution: the fossils still say NO! Institute for Creation Research, El Cajon, California. 391 pp.
Gould, S. J. 1983. Hen's Teeth and Horse's Toes. Norton & Co, New York. 413 pp.
Gould, S. J. & Eldredge, N. 1977. Punctuated equilibrium: the tempo and mode of evolution reconsidered. Paleobiology, 3: 115-151
Gould, S. J. 1991. Bully for Brontosaurus. Penguin, London. 540 pp.
Halstead, L. B. 1984. Evolution - The fossils say yes! In: Montagu, A. (ed.), Science and Creationism. p. 240-254. Oxford University Press, Oxford.
Heilmann, G. 1926. The Origin of Birds. Witherby, London. 208 pp.
Hopson, J. 1980. Relative brain size in dinosaurs. American Association for the Advancement of Science, Selected Symposium, 28: 287-310.
Howgate, M. E. 1984. The teeth of Archaeopteryx and a reinterpretation of the Eichstätt specimen. Zoological Journal of the Linnean Society, 82: 159-175.
Jenkins, F.A. 1993. The evolution of the avian shoulder joint. American Journal of Science, 293A: 253-267.
Jensen, J. A. 1981. Another look at Archaeopteryx as the worlds oldest bird. Encyclia, 58: 109-128.
Jensen, J. A. & Padian, K. 1989. Small pterosaurs and dinosaurs from the Uncompahgre fauna (Brushy Basin Member, Morrison Formation: ?Tithonian), Late Jurassic, western Colorado. Journal of Palaeontology, 63: 364-373.
Jerison, H.J. 1973. The Evolution of the Brain and Intelligence. Academic Press, New York.
Jordan, D. S. & Kellogg, V. L. 1916. Evolution and Animal Life. D. Appleton & Co., New York. 489 pp.
Kitcher, P. 1982. Abusing Science: The Case Against Creationism. MIT Press, Cambridge. 213 pp.
Lowe, P.R. 1935. On the relationships of the Struthiones to the dinosaurs and to the rest of the avian class, with special reference to the position of Archaeopteryx. Ibis, 13: 398-432.
Lull, R. S. 1940. Organic evolution. Macmillan, New York. 743 pp.
McGowan, C. 1984. In the Beginning: A Scientist Shows Why the Creationists are Wrong. Prometheus, Buffalo. 208pp.
Milner, A. C. 1993. Ground rules for early birds. Nature, 362: 589.
Morris, H. 1967. Evolution and the Modern Christian. Presbyterian and Reformed Pub Co, Philadelphia PA.
Newton, A. 1884. Ornithology. In: Encyclopaedia Britannica, 9th. ed. Vol. 18: 2-25.
Ostrom, J. H. 1976. Archaeopteryx and the origin of birds. Biological Journal of the Linnean Society, 8(2): 91-182.
Ostrom, J. H. 1986. The Jurassic "bird" Laopteryx priscus reexamined. Contributions to Geology, Special Paper, 3: 11-19.
Ozawa, T. 1975. Evolution of Lepidolina multiseptata (Permian foraminifera) in East Asia. Memoir of the Faculty of Science, Kyushu University, Series D Geology, 23: 117-164.
Parker, T.J. & Haswell, W.A. 1940. A Text-Book of Zoology. Macmillan and Co., London. 758 pp.
Raup, D. M. 1983. The geological and paleontological arguments of creationism. p. 147-162. In: Godfrey, L. R. (ed), Scientist Confront Creationism. Norton & Co, New York.
Romanoff, A. L. 1960. The Avian Embryo. Structural and Functional Development. Macmillan, New York. 1305 pp.
Strahler, A. N. 1987. Science and Earth History. Prometheus, Buffalo. 552 pp.
Thulborn, R. A. & Hamley, T. L. 1982. The reptilian relationships of Archaeopteryx. Australian Journal of Zoology, 30: 611-634.
Tucker, B. W. 1938. Functional evolutionary morphology: the origin of birds. In: de Beer, G. R. (ed.): Evolution. Essays on Aspects of Evolutionary Biology. p. 321-326. Clarendon Press, Oxford.
Wellnhofer, P. 1993. The seventh specimen of Archaeopteryx from the Solnhofen Limestone. Archaeopteryx, 11: 1-47.
Whetstone, K. N. 1983. Braincase of Mesozoic birds: I. New preparation of the "London" Archaeopteryx. Journal of Vertebrate Paleontology, 2: 439-452.
Whybrow, P. J. 1982. Preparation of the cranium of the holotype of Archaeopteryx lithographica from the collections of the British Museum (Natural History). Neues Jahrbuch für Geologie und Paläontologie, 1982(3) 184-192.