Fossil Hominids: mitochondrial DNA

In July 1997, the first successful extraction of Neandertal DNA was announced. In an article in the journal Cell, a team of German and American researchers led by Svante Pääbo (Krings et al. 1997) claimed to have extracted mitochondrial DNA (mtDNA) from a piece of bone cut from the upper arm of the first recognised Neandertal fossil, the individual found at the Feldhofer grotto in the Neander Valley in Germany in 1856 (Kahn and Gibbons 1997, Ward and Stringer 1997). What is the significance of this study?
What is mitochondrial DNA?
DNA (deoxyribonucleic acid) is the gigantic molecule which is used to encode genetic information for all life on Earth. DNA molecules consists of a long strand of base molecules arranged in the form of a double helix. The bases are adenine, guanine, cytosine, and thymine, often abbreviated as A, G, C, and T. What we ordinarily think of as "our" DNA, because it controls most aspects of our physical appearance, is also known as "nuclear DNA", because every cell in our bodies contains two copies of it in the cell nucleus.
Mitochondria (singular: mitochondrion) are small energy-producing organelles found in cells. Surprisingly, mitochondria have their own DNA molecules, entirely separate from our nuclear DNA. Most cells contain between 500 and 1000 copies of the mtDNA molecule, which makes it a lot easier to find and extract than nuclear DNA. In humans the mtDNA genome consists of about 16,000 base pairs (far shorter than our nuclear DNA), and has been completely sequenced (for one individual, at least; Anderson et al. 1981). What makes mtDNA particularly interesting is that, unlike nuclear DNA which is equally inherited from both father and mother, mtDNA is inherited only from the mother, because all our mitochondria are descended from those in our mother's egg cell (there may be rare exceptions to this rule, however; see below).
Consider the set of all women living today, then the set of all their mothers, and so on. Obviously, each set will be as small as or smaller than the previous set. Eventually the set will contain only one woman, who is known as "mitochondrial Eve". The mtDNA of all living humans is inherited from mitochondrial Eve.
Normally our mtDNA is identical to that of our mother. But, like all DNA, mtDNA mutates occasionally so that one of the bases (A, C, G, or T) changes to a different base. Because of these mutations, human mtDNA has been slowly diverging from the mtDNA of mitochondrial Eve, and the amount of mutation is roughly proportional to the amount of time that has passed. This means that similarity of mtDNA for any two humans provides a rough estimate of how closely they are related through their maternal ancestors. If they have identical mtDNA, they are fairly closely related, maybe even siblings. If they have very different mtDNA, it means their last common maternal ancestor lived long ago.
However, using the genetic difference to estimate the time of the last common ancestor is difficult, for a couple of reasons. One is that the rate at which mtDNA mutates is poorly measured. The other is that even if the average mutation rate is accurately known, some lineages will as a matter of chance accumulate fewer or more mutations than average.
Extraction of the mitochondrial DNA
After death, DNA starts degrading immediately. It is thought that under the most favorable conditions, some DNA fragments can survive for as long as 50,000 to 100,000 years. The Feldhofer Neandertal fossil, thought to be between 30,000 and 100,000 years old, was therefore pushing the limits for this kind of work. However initial testing of the fossil showed good preservation of amino acids, indicating that it might contain recoverable mtDNA.
Polymerase Chain Reaction (PCR) is a technique which can be used to create many copies of an initially small number of molecules. The researchers used PCR to amplify and extract many short strands of mtDNA from the Neandertal sample. By overlapping these, they were able to generate a sequence of 379 bases apparently from the Neandertal individual. To protect against errors and contamination, each base was extracted in at least two separate amplifications. The sequence was extracted from a section of the mtDNA genome known as the hypervariable region 1 (HVR1), so-called because that section of the genome accumulates mutations more rapidly than most of the genome and hence is particularly useful in distinguishing between different populations.
Krings et al. then compared this sequence against a database of 994 different mtDNA sequences from modern humans. For the sequence of mtDNA in question, humans on average differ from each other in 8 +/- 3.1 positions (the '3.1' represents one standard deviation). The greatest difference between any two modern humans was 24, and the smallest difference was 1 (because duplicates were removed from the database).
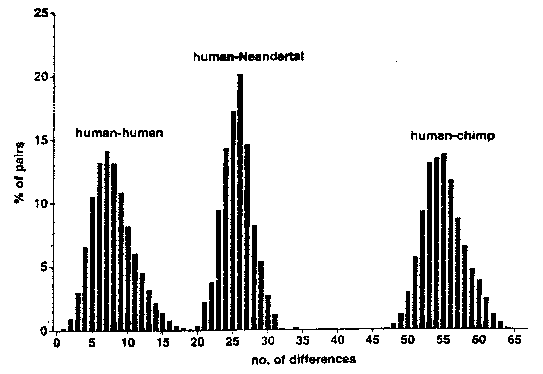
Distributions of Pairwise Sequence Differences among Humans, the Neandertal, and Chimpanzees. X axis, the number of sequence differences; Y axis, the percent of pairwise comparisons. (Krings et al, 1997) |
By contrast, the Neandertal genome had an average of 27 +/- 2.2 differences from modern humans (3.375 times the average difference between modern humans). The smallest difference between any human and the Neandertal was 22, and the largest difference between any human and the Neandertal was 36. These differences put the Neandertal genome well outside the limits of modern humans. Another interesting result is that the mtDNA sequence seemed equally distant from all modern groups of humans. In particular, it did not seem to be more closely related to Europeans, something that might have been expected if, as some scientists think, Neandertals were at least partly ancestral to them.
In 1999, the same workers successfully extracted a second sequence of 340 base pairs of mtDNA from the same Neandertal fossil (Krings et al. 1999). This study confirmed the results of the first. When differences were calculated between the 600 comparable base pairs of 663 modern humans, the Neandertal, and 9 chimpanzees, modern humans differed from each other by 10.9 +/- 5.1 (range 1-35), the Neandertal differed from humans by 35.3 +/- 2.3 (range 29-43), and humans and the two Neandertals both differed from chimps by about 94.
mtDNA from a second Neandertal
In 1999, scientists successfully extracted a 345 base pair sequence of mtDNA from a second Neandertal, a 29,000 year-old fossil of a baby recently discovered in Mesmaiskaya cave in south-western Russia. (Ovchinnikov et al. 2000, Höss 2000) The results of this study were similar to the previous ones, putting the Mezmaiskaya specimen outside the range of modern human mtDNA.
In addition, the two Neandertals are fairly similar, differing from each other in 12 base pairs. The difference is greater than that usually found between pairs of modern Europeans or Asians (only 1% of whom differ in 12 or more places), but comparable to the differences between modern Africans (37% of whom differ by 12 or more).
The distance between Mezmaiskaya and a particular modern human sequence known as the reference sequence (Anderson et al. 1981) was 22, compared to 27 for the first Neandertal. (However, no figures are given for the minimum, average and maximum distances between Mezmaiskaya and modern humans; it is unclear whether Mezmaiskaya is in general closer to modern humans than Feldhofer is.)
The phylogenetic analyses of Ovchinnikov et al. show the two Neandertals grouped together, and separated from all modern humans. As with the first specimen, Mezmaiskaya also appears to be equidistant from all groups of modern humans, strengthening the conclusion that Neandertals are not closely related to modern Europeans.
Because this second individual was discovered about 2,500 km (1,500 miles) from the first, it provides very strong confirmation of the previous results.
The fact that its mtDNA was also fairly close to that of the first Neandertal makes it much less likely that Neandertals and the ancestors of modern humans were both part of an interbreeding population with a large amount of mtDNA genetic variation that has been mostly lost:
"In particular, these data reduce the likelihood that Neanderthals contained enough mtDNA sequence diversity to encompass modern human diversity" (Ovchinnikov et al. 2000)
Interestingly, the preservation of the Mezmaiskaya specimen appears to be much better than that of the Feldhofer specimen. It is so good, in fact, that there is a possibility that its entire mtDNA genome may be able to be sequenced, and there is even a possibility that some of its nuclear DNA may be retrievable.
mtDNA from a third Neandertal
In 2000, scientists announced the sequencing of a third Neandertal mtDNA specimen from a cave at Vindija, Croatia (Krings et al. 2000). When the three Neandertals are compared with modern humans, all three of them cluster together, and apart from all modern humans. This conclusion is reinforced by a study by Knight (2003). Knight excluded from the comparison sites in the mtDNA genome which are known to have mutated more than once, and which are therefore poor indicators of phylogenetic relationships. His study strongly confirmed earlier ones showing deeply divergent histories for modern human mtDNA lineages and the known Neandertal ones.
|
Like modern humans, Neandertals had low genetic diversity compared to apes. The diversity of the three Neandertal mtDNA sequences (3.73%) is lower than that of chimpanzees (14.82+/-5.7%) and gorillas (18.57+/-5.26%), and similar to that of modern humans worldwide (3.43+/-1.22%). If modern humans are sorted into continental groups, the diversity of the three Neandertals is similar to (within one standard deviation of) that for Africans, Asians, native Americans and Australian aboriginals, and Oceanians. Modern Europeans, who live in approximately the same region as the Neandertals, have less diversity than the Neandertals.
Still more Neandertal mtDNA
Schmitz et al. (2002) reported on a fourth Neandertal mtDNA sequence from the second Neandertal fossil found at Feldhofer, the site in Germany at which the first Neandertal fossil was found. It was closely related to the previous Neandertal mtDNA sequences.Serre et al. (2004) were able to sequence mtDNA from four other Neandertal fossils, along with mtDNA from five early modern humans. The four Neandertals all had mtDNA similar to those found in the previous Neandertals. Serre and his colleagues found no evidence of mtDNA gene flow between modern humans and Neandertals in either direction, but could not rule out the possibility of limited gene flow. Interestingly, the mtDNA sequences from the Vindija Neandertals, which have a less extreme Neandertal anatomy than the classic Neandertals, and are considered transitional between modern humans and classic Neandertals by some scientists, were no closer to modern humans than the rest of the Neandertal fossils.
As of 2008, 17 mtDNA sequences have been extracted from Neandertal fossils (Green et al. 2008). All these sequences support the conclusion that Neandertal mtDNA is outside the range of modern human mtDNA, and strongly indicate that Neandertals made no lasting contribution to the human mtDNA gene pool. (This does not prove that they made no contributions to the nuclear DNA of any modern humans, however.)
The first complete Neandertal mtDNA genome
In 2008, the first complete sequencing of Neandertal mtDNA was announced (Green et al. 2008). A complete mtDNA genome of 16,565 base pairs was extracted from a 38,000 year old fossil from the Vindija cave in Croatia. As Krings et al. 2007 had done, the authors created a graph showing the numbers of base pair differences for humans, chimps and the Neandertal when compared against humans. Because they were able to compare across the whole genome rather than a small portion of it, the differences between humans and the Neandertal was far more striking:
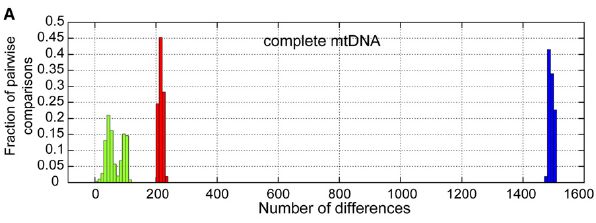
Green: human/human comparisons; Red: human/N'tal comparisons; Blue: human/chimp comparisons. X axis, the number of sequence differences; Y axis, the fraction of pairwise comparisons. (Green et al. 2008) |
Among the humans, the sequences had between 2 and 118 differences. The number of differences between the human mtDNAs and the Neandertal mtDNA varies from 201 to 234. The miniscule overlap between the human figures and the Neandertal figures in the original graph has disappeared and instead there is now a sizeable gap between the human and Neandertal results. Green et al. concluded that:
Analysis of the assembled sequence unequivocally establishes that the Neandertal mtDNA falls outside the variation of extant human mtDNAs, and allows an estimate of the divergence date between the two mtDNA lineages of 660,000 140,000 years.
The first Neandertal nuclear genome
In May 2010, a first draft of a Neandertal nuclear genome was published for the first time (Green et al. 2010). About 4 billion base pairs, or roughly 2/3rds of of the entire genome, was sequenced from three individuals. Even more interestingly, analysis of the genome seems to show that Neandertals interbred with humans, and that all non-African modern humans contain between 1% and 4% of Neandertal genes. (A later paper, Reich et al. 2010, has narrowed that range to between 1.9% and 3.1% of Neandertal genes.) Because Asians as well as Europeans have these Neanderthal genes, the researchers believe the most likely explanation is that the interbreeding occurred in the Middle East when modern humans first left Africa between 60,000 and 80,000 years ago, and before they expanded into the rest of the world.Is the Neandertal mtDNA outside the human range?
Yes.
Note that (in reference to Krings et al. 2007) because two modern human sequences are 24 bases apart, while the smallest Neandertal/human difference is only 22, this does not mean that the Neandertal sequence is within the range of modern humans. To use an analogy, suppose we measured the height of 994 adult humans, and they varied from 4'8" to 6'8" (a difference of 24 inches). Suppose we then found a skeleton which was 8'6" in height. No one would claim that it fell within the modern human range because it was closer to the nearest human (22 inches) than the tallest human was from the shortest human (24 inches).
Note also that the two figures (22 and 24) are measuring very different things, making it invalid to compare the two figures. Just as the Neandertal was compared against 994 modern humans, any of those humans could be similarly compared against the other 993 humans. We could compute the minimum, average, and maximum distance from that human to the other humans, just as was done for the Neandertal. If we calculated those values for all the humans, we could then calculate minimum, average and maximum values of all the individual minima, averages and maxima, and compare those values against the equivalent values for the Neandertal.
We do not know from the Krings et al. 1997 paper the distribution of minimum distances of humans from other humans. The smallest such value is 1. The largest such value might, I suspect, be as much as 5. The same value for the Neandertal is 22, well outside the human range.
For average distances of humans to other humans, we know the average value is 8.0. The minimum average distance will be a little less; the maximum average value must be at least 12 (this can be deduced from the fact that there are two humans 24 apart) and less than 24; I would guess it might be about 16 for a highly atypical human. For the Neandertal, the value is 27, again well outside the human range.
For maximum distances, the maximum such value is 24, but for most humans, the maximum distance to any other human will be less than that. The value of 24 is highly atypical, because it is taken between the two individuals who have indepently diverged farthest from mitochondrial Eve, and is the maximum of nearly half a million (994 * 993 / 2) comparisons among modern humans. For the Neandertal, the value is 36, again well outside the human range.
In other words, for all three measurements (minimum, average and maximum distances to other humans), the Neandertal measurement is much larger than the maximum value of the same measurement from a sample of 994 modern humans, and even further from the average value. The Neandertal is not merely outside the human range, but well outside it.
The extraction of a complete Neandertal mtDNA genome (Green et al. 2008) makes the above discussion moot, because we now know that when the full genome is considered, the imaginary overlap between human-human and human-Neandertal differences that creationists misrepresented entirely disappears.
Possible problems
The use of mitochondrial mtDNA to investigate human history is not without drawbacks.The rate of mtDNA mutation is not well known. A study by Parsons et al. (1997) found a rate 20 times higher than that calculated from other sources. In an article reviewing mtDNA research, Strauss (1999a) reports that mtDNA mutation rates differ in some groups of animals, and can even vary dramatically in single lineages. Although there are many agreements, some divergence dates for modern animals calculated from mtDNA do not match with what is known from the fossil record. There are suggestions from a few sources that paternal mtDNA can sometimes be inherited, which could affect analyses based on mtDNA.
In 1999 Awadalla et al. published a study suggesting that mtDNA could sometimes be inherited from fathers. If mtDNA is inherited only from mothers, the correlation between different mutations should not depend on how far apart on the genome they were. Instead, their measurements showed that mutations at distant sites on the mtDNA genome were less likely to be correlated than nearby mutations, suggesting that mtDNA from mothers and fathers could sometimes get mixed. However, there is no explanation so far as to how this recombination could be occurring, and the possibility that other phenomena could be causing this effect has not yet been disproved. If it occurs, mixing would mean that the dates from current mtDNA studies would be too old. If mixing is common enough, it could even mean that there was no mitochondrial Eve, because different parts of the mtDNA molecule would have different histories. (Awadalla et al. 1999, Strauss 1999b) Other studies, however, have contradicted these results and argued for strictly maternal mtDNA inheritance (Elson et al. 2001), and, according to Sykes (2001), the Awadalla paper and another paper which also suggested that mtDNA could be inherited paternally were based on incorrect data and were later retracted.
Conclusions
The studies of Neandertal mtDNA do not show that Neandertals did not or could not interbreed with modern humans. But the lack of diversity in Neandertal mtDNA sequences, combined with the large differences between Neandertal and modern human mtDNA, strongly suggests that Neandertals and modern humans developed separately, and did not form part of a single large interbreeding population. However Neandertals apparently remained capable of interbreeding with humans, and did so with an early population of modern humans in the Middle East about 70,000 years ago.References
Anderson S., Bankier A.T., Barrel B.G., de Bruijn M.H.L., Coulson A.R., Drouin J. et al. (1981): Sequence and organization of the human mitochondrial genome. Nature, 290:457-74.
Awadalla P., Eyre-Walker A., and Smith J.M. (1999): Linkage disequilibrium and recombination in hominid mitochondrial DNA. Science, 286:2524-5.
Elson J.L., Andrews R.M., Chinnery P.F., Lightowlers R.N., Turnbull D.M., and Howell N. (2001): Analysis of European mtDNAs for recombination. American Journal of Human Genetics, 68:145-53.
Green R., Malaspinas A-S, Krause J., Briggs A., et al. (2008) A Complete Neandertal Mitochondrial Genome Sequence Determined by High-Throughput Sequencing. Cell, 134:416-426.
Green et al. 2010: A draft sequence of the Neanderthal genome. Science, 328:710.
Höss M. (2000): Neanderthal population genetics. Nature, 404:453-4.
Kahn P. and Gibbons A. (1997): DNA from an extinct human. Science, 277:176-8.
Knight A. (2003): The phylogenetic relationship of Neandertal and modern human mitochondrial DNAs based on informative nucleotide sites. Journal of Human Evolution, 44:627-32.
Krings M., Capelli C., Tschentscher F., Geisert H., Meyer S., von Haeseler A. et al. (2000): A view of Neandertal genetic diversity. Nature Genetics, 26:144-6.
Krings M., Geisert H., Schmitz R.W., Krainitzki H., and Pääbo S. (1999): DNA sequence of the mitochondrial hypervariable region II from the Neandertal type specimen. Proceedings of the National Academy of Sciences, USA, 96:5581-5.
Krings M., Stone A., Schmitz R.W., Krainitzki H., Stoneking M., and Pääbo S. (1997): Neandertal DNA sequences and the origin of modern humans. Cell, 90:19-30.
Ovchinnikov I.V., Götherström A., Romanova G.P., Kharitonov V.M., Lidén K., and Goodwin W. (2000): Molecular analysis of Neanderthal DNA from the northern Caucasus. Nature, 404:490-3.
Parsons T.J., Muniec D.S., Sullivan K., Woodyatt N., Alliston-Greiner R., Wilson M.R. et al. (1997): A high observed substitution rate in the human mitochondrial DNA control region. Nature Genetics, 15:363-8.
Reich, Green, Kircher et al. 2010: Genetic history of an archaic hominin group from Denisova Cave in Siberia. Nature 468:1053.
Schmitz R.W., Serre D., Bonani G., Feine S., Hillgruber F., Krainitzki H. et al. (2002): The Neandertal type site revisited: Interdisciplinary investigations of skeletal remains from the Neander Valley, Germany. Proceedings of the National Academy of Sciences, USA, 99:13342-7.
Serre D., Langaney A., Chech M., Teschler-Nicola M., Paunovic M., Mennecier P. et al. (2004): No evidence of Neandertal mtDNA contribution to early modern humans. PLoS Biology, 2:313-7.
Strauss E. (1999a): Can mitochondrial clocks keep time? Science, 283:1435
Strauss E. (1999b): mtDNA shows signs of paternal influence. Science, 286:2436
Sykes B. (2001): The seven daughters of Eve. London: Bantam Press. (a good popular book about mitochondrial DNA)
Ward R. and Stringer C.B. (1997): A molecular handle on the Neanderthals. Nature, 388:225-6.
Commentaries
Nature, feature of the week, March 30, 2000: Neanderthal DNANeandertal mitochrondrial sequence stored at GenBank (accession number AF011222)
DNA Shows Neandertals Were Not Our Ancestors (Pennsylvania State University)
DNA clues to Neanderthals: mtDNA from a third Neandertal
Neandertal DNA, by Mark Rose (Archaeological Institute of America)
Other references about mitochondrial DNA
What, if anything, is a Mitochondrial Eve?, by Krishna KunchithapadamThe Mitochondrial DNA concordance, by Kevin Miller and John Dawson
The Fire Within: The Unfolding Story of Human Mitochondrial DNA, by Ken Miller
Oxford Ancestors: trace your mtDNA lineage
This page is part of the Fossil Hominids FAQ at the talk.origins Archive.
Home Page |
Species |
Fossils |
Creationism |
Reading |
References
Illustrations |
What's New |
Feedback |
Search |
Links |
Fiction
http://www.talkorigins.org/faqs/homs/mtDNA.html, 22/05/2011
Copyright © Jim Foley
|| Email me 