Other Links:
|
The story of Haeckel's embryos is different in an important way from that of the other chapters in Jonathan Wells' book. As the other authors show, Wells has distorted ideas that are fundamentally true in order to make his point: all his rhetoric to the contrary, Archaeopteryx is a transitional fossil, peppered moths and Darwin's finches do tell us significant things about evolution, four-winged flies do tell us significant things about developmental pathways, and so forth. In those parts of the book, Wells has to try and cover up a truth by misconstruing and misrepresenting it.
In the case of Haeckel, though, I have to begin by admitting that Wells has got the core of the story right. Haeckel was wrong. His theory was invalid, some of his drawings were faked, and he willfully over-interpreted the data to prop up a false thesis. Furthermore, he was influential, both in the sciences and the popular press; his theory still gets echoed in the latter today. Wells is also correct in criticizing textbook authors for perpetuating Haeckel's infamous diagram without commenting on its inaccuracies or the way it was misused to support a falsified theory.
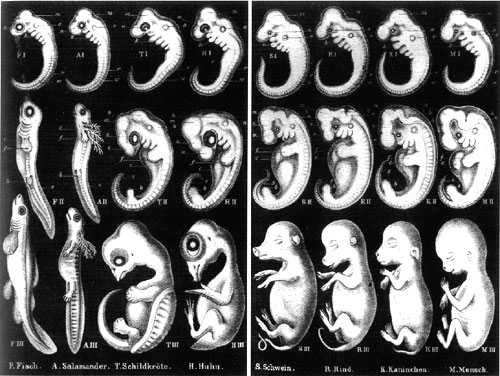
Unfortunately, what Wells tries to do in this chapter is to take this invalid, discredited theory and tar modern (and even not so modern) evolutionary biology with it. The biogenetic law is not Darwinism or neo-Darwinism, however. It is not part of any modern evolutionary theory. Wells is carrying out a bait-and-switch here, marshalling the evidence and citations that properly demolish the Haeckelian dogma, and then claiming that this is part of "our best evidence for Darwin's theory."
Who was Haeckel, and what was his theory?
Ernst Haeckel (1834-1919) was an extremely influential German scientist. He had an enviable reputation as a comparative embryologist, but his primary claim to fame was that he was an early adopter of Darwin's theory who used the evidence of embryology to support evolution. Wells is quite correct to mention the importance of Haeckel in 19th century biology. There is no doubt that his efforts to popularize the theory were important in giving evolution credibility in the scientific establishment, and to laymen as well. Darwin and Haeckel met and corresponded, and each influenced the theories of the other strongly. However, Haeckel's theories owed an even greater debt to an earlier philosophical tradition, in particular the work of Goethe and Lamarck. Although Darwin was appreciative of Haeckel's support for natural selection, he was also tentative in using Haeckel's ideas in his writings; Darwin relied far more on von Baer's embryological data to support common descent.
Furthermore, Haeckel's theory was rotten at the core. It was wrong both in principle and in the set of biased and manipulated observations used to prop it up. This was a tragedy for science, because it set evolutionary biologists and developmental biologists down a dead-end, leading to an unfortunate divorce between the fields of development and evolution that has only recently been corrected.
Haeckel's theory is encapsulated in his memorable aphorism, "ontogeny recapitulates phylogeny," also called the biogenetic law. What that means is that development (ontogeny) repeats the evolutionary history (phylogeny) of the organism - that if we evolved from a fish that evolved into a reptile that evolved into us, our embryos physically echo that history, passing through a fish-like stage and then into a reptile-like stage. How could this happen? He argued that evolutionary history was literally the driving force behind development, and that the experiences of our ancestors were physically written into our hereditary material. This was a logical extension of his belief in Lamarckian inheritance, or the inheritance of acquired characters. If the activity of an organism can be imprinted on its genetics, then development could just be a synopsis of the activities of the parents and grandparents and ever more remote ancestors. This was an extremely attractive idea to scientists; it's as if development were a time machine that allowed them to look back into the distant past, just by studying early stages of development.
Unfortunately, it was also completely wrong.
The discoveries that ultimately demolished the underlying premises of the biogenetic law were the principles of genetics and empirical observations of embryos. Lamarckian inheritance simply does not occur, as we all learned in middle school biology. DNA is the agent of heredity, and it is not modified by our ordinary actions - if you should get a tattoo, it is not also written into the chromosomes of your sperm or ova, and there's no risk that your children will be born with "Mom" etched on their arm. The discovery that Haeckel had taken unforgivable shortcuts with his illustrations was a relatively minor problem for his theory, because the general thrust of his observations (that vertebrate embryos resemble each other strongly) had been independently confirmed. What really scuttled the whole theory was that its foundation was removed, which makes the claim by Wells that biologists are trying to "resurrect recapitulation" rather peculiar, since there aren't very many who see any validity to Lamarckian heredity.
Wells would also like the reader to think that Haeckel's embryos were a key piece of the puzzle that Darwin himself used to assemble his theory, and that therefore Darwinism was built on a false foundation. He neglects to mention an important matter of chronology, however. Darwin's Origin of Species was published in 1859, after decades of research. Haeckel's theory was first published in 1866 , and Haeckel's diagrams were published in 1874. Haeckel did not have any significant input into the Origin, but Darwin (and Lamarck and a number of German philosophers, such as Goethe) did profoundly influence Haeckel. Clearly, the later elaborations by Darwin on the theory of evolution were complemented by Haeckel's ideas, in particular his ideas about heredity, but those are precisely the components of Darwinism that were invalidated by Mendelian genetics and the neo-Darwinian synthesis!
What is the phylotypic stage?
Darwin said, "Hardly any point gave me so much satisfaction when I was at work on the Origin as the explanation of the wide difference in many classes between the embryo and the adult animal, and the close resemblance of the embryos within the same class." Early in the 19th century, von Baer noted that the embryos of different species could not be as easily distinguished from one another as are the adults. This is a simple observation that has been made numerous times in the past few hundred years, an observation that is not tied to any particular theory, whether of creation or evolution -- von Baer himself made these observations 30 years before Darwin published, and did not accept evolution then or later. That is, while vertebrate adults may look very different from one another, vertebrate embryos all go through a period in development in which they all resemble each other more strongly. This period is called the phylotypic stage. At this time in development, vertebrate embryos all express a suite of characters that are common to the entire vertebrate lineage: they have a notochord and a dorsal nerve cord, they have pharyngeal arches and a tail, and they have a repeating series of blocks of muscle called somites. Most of the features that distinguish different vertebrate groups, such as limbs or fins, hair or feathers or scales, and wings or forearms, haven't yet developed. In addition, we now know that this is the period during which a set of crucial pattern forming genes (the emx/otx/Hox genes) is first expressed, and lay down the molecular blueprint of the body plan.
The observation is not tied to any particular theory, but we still have to explain the existence of this phenomenon in any theory of origins. Haeckel tried. His explanation was a flawed model of heredity that claimed that there were constraints on the properties of genetics, such that only the final phases were easily modified. This meant that new features were usually added only at the end of the program of development, so that the sequence of development was roughly the same as the sequence of evolution. However, this explanation has been falsified. Not only does genetics not work that way, but development does not proceed in a fashion that resembles the evolutionary sequence; as Wells points out (echoing the same criticisms that have been made for over a hundred years), the very earliest stages of development can differ radically. They then converge on a similar appearance at the phylotypic stage, and diverge in morphology again.
Modern theories of development and evolution propose something that fits the observations, and that Wells cannot easily dismiss. Genes can be modified to act at virtually any point in development, so the theoretical constraint imposed by Haeckel is nonexistent. Variations between species at the earliest stages were a problem for Haeckel, but are not incompatible at all with modern developmental biology. There isn't even a requirement for absolute morphological identity at the phylotypic stage. As Wells points out, Michael Richardson has been identifying variation within that stage between species.
In Icons of Evolution, Wells is flogging a dead horse by exclusively bashing Haeckel's explanation, and ignoring modern ideas. Furthermore, he is not making any contribution to our understanding of why vertebrates develop the way that they do. Why should animals that differ in appearance as adults exhibit similarities as embryos? Why should human embryos have tails and build their faces from the same embryonic foundation that fish use to build gills? These are questions that Wells fails to address, and his approach seems to be to try to argue instead that they do not exhibit similarities as embryos. Yet they do. These similarities argue for common ancestry.
What about the early differences in development?
Wells makes much of the fact that the biogenetic law presumes that embryos ought to be most similar at the earliest possible stage of development, yet the period of greatest similarity, the phylotypic stage, occurs well into development, and that the earliest stages exhibit seemingly significant differences. This is true, but irrelevant.
- The biogenetic law is not Darwinism. Poking holes in the biogenetic law is easy to do, it's been done repeatedly in the scientific literature, and it does not damage modern evolutionary biology at all.
- This observation isn't news. Von Baer pointed out in the 1820's that the earliest tissues to differentiate in the mammalian embryo are the extraembryonic membranes, which also happen to be among the more recent features in the mammalian lineage to evolve. The figure Wells uses to make this point (Figure 5-3) is a redrawn version of a similar figure published by Elinson in 1987.
- Wells downplays the similarities that exist to emphasize the differences. This is the same biased game that Haeckel played: the only difference is that he emphasized the similarities rather than the differences.
That last point requires further explanation. What modern developmental biology has been discovering is that these earliest stages in different vertebrate embryos are substantially similar. The differences are superficial, and in many cases a consequence of different amounts of maternal investment. For instance, one of the first tasks of a fertilized, single-celled egg is to divide repeatedly to produce a large number of cells. In animals that have a large amount of yolk, such as a fish, reptile, or bird, the yolk is too large to be efficiently divided, so the cell divides partially, producing a cap of cells on top of the yolk. In animals that have relatively little, well-dispersed yolk, such as the frog, or have no yolk at all, as in mammals, the egg can divide completely, and produces a ball of cells. That's the extent of the difference, whether you have a mass of cells by itself or a mass sprawled on top of a yolk.
These differences in shape lead to some apparent differences in the next step of development, gastrulation. The critical function of gastrulation is that some cells have to move inward, into the mass of cells. This movement of sheets of cells generates the three-dimensional structure of the embryo, and during the process, gives cells opportunities to interact and induce new tissue types. Whether the embryo is a ball of cells or a mass on top of a yolk, though, all vertebrates carry out equivalent movements during gastrulation; again, the differences are superficial, depending on whether the cluster of cells is balled up or flattened. There are other differences that can be cataloged, such as whether the cells move inward as a coherent, adhesive sheet or whether they delaminate and move in as single cells, but again, the core process, that they move inward to set up new layers, is the same.
On a more fundamental level, it appears that vertebrates share a common set of signaling molecules, such as BMP-4 and chordin, that are important in defining polarity and specifying tissue types at these early stages. The processes of cell signaling important in inducing new tissues during gastrulation are similar in all vertebrates, and the same answers are turning up in fish and mice, despite the morphological differences in their layouts. It is misleading to focus on apparent differences in shape on a gross level, when the deeper, molecular aspects of these stages of development are homologous.
What's wrong with "gill slits"?
Wells is quite unhappy with the common term "gill slits." He spends several pages telling us that embryonic mammals don't have gills, and that even at the stages that fish have gill slits, they don't have gills. He is missing the point, however: nobody has claimed that they do have gills. They have "gill slits," a completely different thing altogether. He has gotten so wrapped up in a trivial etymological argument that he has lost all sight of biological reality.
Vertebrate embryos universally have prominent structures in their neck region that are called by various names in the scientific literature: branchial, pharyngeal, or visceral pouches or grooves or furrows or arches. Because they may appear as a repeating series of slits in the neck of the embryo, resembling the pattern of repeated elements in the neck of adult fish, they have also been colloquially called "gill slits" or "gill pouches." They are not, however, gills - and scientists have not been claiming that they are (Wells even quotes several authors, Wolpert and Rager, who explicitly state this simple, obvious fact). So what are they?
"Gill slits" are common structural elements of vertebrate craniofacial development. "Common" is the important term here. It turns out that all vertebrates build their face in the same, somewhat improbable and counterintuitive way; it is this deep similarity that is the root of the evolutionary argument that it reflects common ancestry.
The head of all vertebrate embryos, whether they are a fish or a human, can be simply described as a curved tube largely made up of presumptive brain (Figure 2), with a series of 4 to 7 finger-like tissues hanging down from it, the pharyngeal arches. What we consider a face, everything from just below the eyes, back to the ears, and down to the neck, is absent. Instead, we have these dangling blobs, each of which will contain a cartilaginous rod, a column of muscle, a significant branch of the circulatory system, and an assortment of other cell types. These arches are reiterated modules that will subsequently merge and rearrange themselves (along with other cranial tissues, most importantly a migrating population of cells from the top of the head called the neural crest) to form the more familiar face. They do so in similar ways in all vertebrates: the first pharyngeal arch, for instance, always forms the jaw, and the second arch always forms the hyoid. There are also differences that emerge in different classes. Pieces of the first two arches find their way into bones of the mammalian ear. The third and subsequent arches in fish end up in the gills, while those same arches in a human form a series of cartilages in the throat. The third fuses with the hyoid, the fourth forms a major part of the thyroid cartilage, and the fifth forms the cricoid and arytenoid cartilages. Non-cartilaginous elements of these structures end up incorporated into all kinds of tissues, glands and muscles and epithelia, of the neck and face.
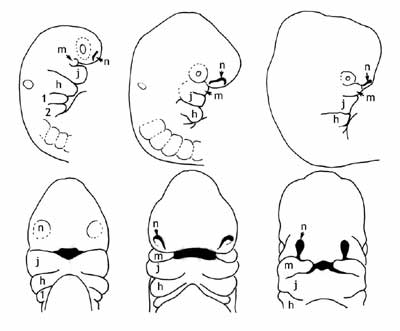 |
This common pattern of development is a genuinely remarkable thing. It is one of the reasons that early evolutionary biologists, such as Charles Darwin, argued that the evidence of embryology was so important to his theory - we see such intricate processes redone over and over again in such similar ways in species after species. Given that the initial pharyngeal arches are radically rearranged over the course of development, there is no obvious reason why all vertebrate embryos begin with virtually identical structures that are equally remote from their final morphology, other than that they reflect a shared morphological foundation and a common ancestry.
Wells wants to deny all this. To do so, he focuses entirely on the terminology. He points out that "gill slits" are not gills, a fact that is well understood by every developmental biologist. He says, "they are never 'gill-like' except in the superficial sense that they form a series of parallel lines in the neck region." This is true, and no one claims any deeper significance to the term than that. However, Wells then tries to claim that "The only way to see 'gill-like' structures in human embryos is to read evolution into development" and that "there is no way 'gill-slits' in human embryos can logically serve as evidence for evolution." These statements are false and misleading. As he himself has pointed out, one way to see "gill-like" structures in embryos is to see that they form a repeated pattern of lines in the neck - that is not presuming evolution, and is a legitimate source for the name. Of course, then we have to wonder why human embryos have these strange lines, and why all other vertebrates also have them...and this commonalty of structure is the evidence for evolution, not the superficial implications of the names.
All vertebrate embryos have pharyngeal arches. Because sometimes these structures are referred to by the simpler term "gill slits" in textbooks does not mean that they do not exist or that the homology disappears, although that is the game Wells would like to play.
Where are all the recapitulationists?
One premise that Wells brings up with regularity in this chapter is that evolutionary biologists have relied on the false doctrines of Haeckel to prop up Darwinian dogma. He claims that Haeckelian theories "have periodically risen, phoenix-like, from the ashes of empirical disconfirmation" throughout the 20th century. He uses section headings like "Resurrecting recapitulation" and "Haeckel is dead. Long live Haeckel." He accuses biologists of a conspiracy of silence, hiding the flaws in Haeckel's work on one hand, and using it as evidence for evolution on the other.
None of this is true.
This is so patently obvious that we only need to use Wells' own scholarship to show it. He cites a number of authors who discuss Haeckel or the biogenetic law:
- Adam Sedgwick, 1894
- William Garstang, 1922
- Gavin de Beer, 1958
- William Ballard, 1976
- Stephen J. Gould, 1977
- Richard Elinson, 1987
- Jane Oppenheimer, 1987
- Michael Richardson, 1995
- Stephen J. Gould, 2000
However, here is the surprising thing: all of these authors condemn the idea that embryonic development follows the evolutionary pattern in no uncertain terms! Sedgwick, for instance, compiled an extensive list of objections to recapitulation as formulated by von Baer and Haeckel, and specifically rejected it as untenable - in 1894. This represents over a century of unambiguous denial of recapitulation. The date can be pushed back even further, since von Baer published his critique of recapitulatory interpretations of his observations in 1828. Wells cites these authors to back up his argument that Haeckel's ideas are false, but at the same time he tries to pretend that evolutionary biologists are all closet fans of Haeckel. If that is the case, where are the citations to prominent modern scientists defending his theories?
The case of Stephen J. Gould illustrates Wells' two-faced strategy particularly well. In 1977, Gould wrote an excellent scholarly book on the intertwined history of embryology and evolution, titled Ontogeny and Phylogeny. As might be guessed from the title, Haeckel is a prominent character in the book, and his theories and their consequences in the field are dissected in detail and without mercy. Gould also returned to this topic in his column in Natural History magazine in 2000, centering his commentary on the discovery of a scathing critique of Haeckel by one of his contemporaries, Louis Agassiz. Gould has also written other related articles, disparaging textbook authors for their deplorable habit of recycling text and figures well beyond reason. Yet how does Wells twist the facts? He claims that Gould "...(despite having known the truth for over twenty years) kept his mouth shut until a "creationist" (actually, a fellow biologist) [he is referring to Michael Behe here] exposed the problem"!
This is a remarkable example of the kind of dishonest doublespeak used by Wells throughout his book. He can cite author after author clearly condemning Haeckel's ideas between 1894 and 2000, and simultaneously claim that biologists have been "resurrecting recapitulation" throughout the 20th century. He claims that Behe "exposed the problem", but Behe has done no research in embryology, has published nothing original on the history of 19th century science, and deserves no credit here - this "problem" was exposed over a century ago, and the real creative and innovative work in embryological analysis being done here is by an evolutionist, Michael Richardson, not Behe. I'm also surprised that Gould can publish an almost 500 page book that is critically well-received and frequently cited, and still be accused of keeping his mouth shut on the subject!
Gould is not alone. The author of one of the most influential textbooks in developmental biology, Scott Gilbert, described the biogenetic law as "disastrous" and "untenable". Raff and Kaufman made the same case against Haeckel that Wells does here, so what Wells is teaching us is not surprising anyone.
Are the textbooks really that bad?
Wells is particularly incensed at the authors of introductory textbooks who, he claims, are misleading their students. I agree that he can reasonably argue that textbooks should not use the obsolete and inaccurate drawings done after Haeckel's work, but, in what I consider the most amusing line in this entire chapter, Wells expresses indignation that "Some textbooks, instead of reproducing or redrawing Haeckel's embryos, use actual photos." How dare those nefarious textbook authors use photographic data to support their ideas!
In his first appendix, Wells rates a number of college textbooks, giving them letter grades for how well their ideas fit with his. All the books get either a D or an F for their handling of vertebrate embryos. However, it is obvious that he is stacking the deck against them.
To get an F, a textbook needs only to use a version of Haeckel's drawings, claim that early similarities in vertebrate embryos are evidence for common ancestry, and refer to gill slits. I agree that books should not use Haeckel's illustrations, unless it is to make a historical point. However, similarities in structure are evidence for common ancestry; it's a little unfair to penalize a book for making a valid point. Similarly, vertebrate embryos do have "gill slits", even if the associations of that term confuse Dr. Wells.
To improve their grade to a D, a book uses "misleading photographs" of real embryos instead of the Haeckelian drawings. Again, however, what is the objection here? These aren't fudged or inaccurate drawings. They are photographs of vertebrate embryos that accurately illustrate what they look like, that anyone with a microscope and access to embryos can see (Figure 3). Is it next on his agenda to condemn classes that allow students to examine live or preserved embryos in the laboratory?
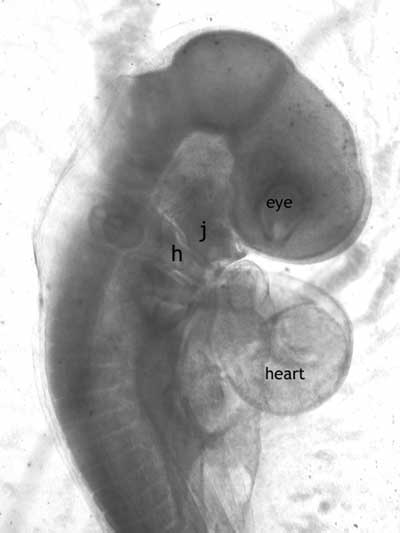 |
What does a textbook have to do to get an A? First, it may not use misleading drawings or photos. Wells has not shown that the photos used in any of these books are misleading at all. Second, it should not use the term "gill slit". This is a capricious requirement; there is nothing categorically wrong with that particular term.
Third, it should discuss dissimilarities at the earliest stages (which is a good idea, but may be impractical in an introductory text) and call this a problem for Darwinian evolution. There is a difficulty in meeting this last requirement, however. The differences may be a problem for a Haeckelian model of evolution, but they are not a problem for modern formulations of evolution. A book that suggested that the early differences are a problem would be straining to misinterpret the evidence.
Finally, his fourth requirement is that the book must consider Darwin's theory of vertebrate origins to be wrong. I presume by this that it must use observations of vertebrate embryos to somehow argue that different vertebrates did not evolve from a common ancestor. Unfortunately, this conclusion does not follow from the evidence. The deep similarities in the process of development for all vertebrates support the idea that they are part of a common lineage; furthermore, progress in the molecular biology of development has been steadily confirming even greater homologies-it is simply not an option to even suggest that these organisms did not have a common ancestor.
One of the textbooks which Wells gives a "D" grade is one I've taught from for a number of years, Campbell's Biology. I took a close look at the relevant section of this particular book. I immediately found one serious shortcoming: this section is all of two paragraphs long, with one figure. It would be good to see more substance on this subject, especially in a book that is 1175 pages long, but one recurrent problem in the books that Wells has targeted is that these are surveys of all of biology, from biochemistry to ecology, so it isn't too surprising that everything gets less depth of treatment than we would like. However, I found nothing in what Campbell has written which is objectionable. He begins with the point that "Closely related organisms go through similar stages in their embryonic development", and illustrates that with a photograph of an avian and mammalian embryo. This statement is correct, and the figure backs up the point. He ends the section by explicitly correcting Haeckel's ideas, saying that "The theory of recapitulation is an overstatement. Although vertebrates share many features of embryonic development, it is not as though a mammal first goes through a 'fish stage', then an 'amphibian stage', and so on. Ontogeny can provide clues to phylogeny, but it is important to remember that all stages of development may become modified over the course of evolution." This is entirely correct. I do not see any errors of fact in Campbell's treatment of the subject, although I do think it is unfortunate that so little space can be spared for it.
This is not to suggest that textbooks are without flaw. I agree with Wells that the contortions by Balinski in his 1975 text to rationalize the biogenetic law are deplorable. I also concur that textbooks that are already giving short shrift to this particular subject ought not to be wasting space with Haeckel's illustration, which is not good biological data and is only of historical interest. Wells goes too far, however. He tries to use one old (if influential) distortion to discard a host of perfectly valid observations in his attempts to discredit some logical conclusions that conflict with his biases.
What lesson should we learn from embryos?
Wells is sending the wrong message in his discussion of Haeckel's embryos. He would like his readers to think that biology has been finding growing disparities in the principles of development, disparities that challenge evolution by casting doubt on the unity of the vertebrate lineage. Nothing could be further from the truth. Molecular biology, in particular, has been showing us that there is a greater degree of similarity between species than previously suspected.
The Hox genes, for example, illustrate this relationship. These are a set of genes that are recognizable by a number of features: they all share a common 180 base pair sequence called the homeobox (from which their name is derived), they are all DNA binding proteins that act to turn other genes on or off, and they have a unique organization. They are all laid out in an order on the chromosome that matches the order of their expression: that is, a Hox gene that is turned on in the head is at one end of the cluster of genes, a Hox gene that is turned on in the tail is at the other end, and all the genes in between are in corresponding spatial order (Figure 4). These genes are involved in patterning the organism, laying down the general body plan. They work to specify where the head should form, for instance, or the neck or lumbar region. Interestingly, these genes are first activated at the phylotypic stage. One good reason for the conservation of the morphology at the phylotypic stage is that this is when these genes have to interact with one another and other genes to sketch out the blueprint for subsequent development. There are global interactions between genes, and genes may be reused in several places. For example, several tissues have been found to induce lens tissue, including heart mesoderm, and changing the timing of heart differentiation could thereby impact eye formation. The specific morphology of the phylotype really does represent a literal foundation upon which the rest of development proceeds, and is resistant to evolutionary change, because there are too many later events that are dependent on it.
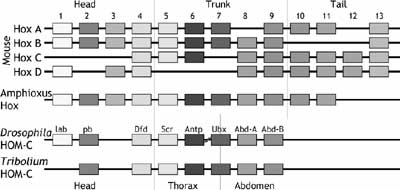 |
All vertebrate embryos use these Hox genes to define their body plan. One surprising result of molecular approaches to development is that it seems that all multicellular animals, everything from worms to insects to mammals to jellyfish, use this same spatially organized array of homeobox-containing genes to define their layout. There are differences: vertebrates seem to have duplicated the array a few times, so that they have four copies of the set of Hox genes, while Drosophila (in which these genes were first identified) has a single cluster; mice and humans have 13 Hox genes in each cluster, while Drosophila has 9, and Hydra has 2. The universality of these genes in animal development has led to the concept of the zootype, or form that links all animals, rather than just the embryos of a single phylum. While Jonathan Wells would like to argue that there is a lack of correspondence between embryos within just the vertebrate lineage, molecular biologists have been discovering information that broadens the correspondence, finding fundamental similarities that link all animals.
Conclusion
Jonathan Wells would like to discredit evolution, and in Haeckel's embryos, he has found a story to his liking. There is a bit of intentional fakery to it, there is a clear affiliation with Darwin himself, and there is a long history of recognition of Haeckel's influence intermingled with unambiguous repudiation of his ideas. All he has to do is try to entangle Haeckel's discredited theories and poor modern reputation with the set of valid observations and modern explanations, and he can bury the truth under innuendo and association. However, we just have to remember three things:
- Evolutionary theory is not founded on Haeckel's observations or theories. Haeckel's work was discredited in the 19 th century, and has not been relevant to biology since the rediscovery of Mendel's laws of genetics. That the biogenetic law is false has been the consensus of biologists for over 100 years, and developmental biologists have been working constructively to provide alternative explanations, which have so far all been evolutionary in nature.
- The similarities between vertebrate embryos are real. We must distinguish between observations of those similarities and hypotheses about their causes. The similarities are not in doubt; there are worthwhile studies of the degree and timing of the similarities, but none that question their overall existence. What Wells has described is one hypothesis about the cause, Haeckel's biogenetic law, which failed early and spectacularly. He has not addressed any modern hypotheses, nor has he provided a better alternative.
- Evidence for common descent lies in the unity of form and process. We do not use Haeckel's outmoded, invalid mechanism to argue for evolution. Instead, we look at the marvelous convergence of disparate organisms on common principles: all animals use the same genes to define regions of their bodies, all vertebrates build their faces by unlikely rearrangements of odd pharyngeal protrusions, and even tailless mammals like us have to start with tailed embryos. The best explanation for these phenomena is that they are a consequence of a common heritage.